Jesús Pineda
Biology Department MS 34
Woods Hole Oceanographic Institution
Woods Hole, MA 02543
jpineda@whoi.edu
(508) 289-2274
The problem of larval return for nearshore species.
The majority of species associated to the seafloor inhabiting the nearshore, ~0 to 30 m water depth, have a two-phase life cycle, where adults are conspicuous and live associated to the seafloor, while larvae are minuscule, and drift at the mercy of the currents. After a period of time ranging from minutes to years, with a ~1 month mode for temperate species, larvae drift, diffuse, and accumulate in an offshore larval pool, far from their adult habitats. These larvae must return shoreward in order to complete their life cycle. Transport of larvae from offshore to nearshore waters is critical for understanding issues from population dynamics to species geographical ranges, from fisheries-management to design of marine reserves.
This problem involves larval behavior and physical transport mechanisms.
Invertebrate larvae are generally small (~ < 1 mm), with limited swimming
capabilities, and it is generally agreed that because larval swimming speeds
are so small, these larvae must transport shoreward by some sort of advection.
On the other hand, by controlling its vertical position within the water
column, larvae may exploit different transporting or concentrating mechanisms.
Understanding physical transport for nearshore species involves characterizing
the mechanism and its temporal and spatial variability. One must understand
how transport occurs, and also the secondary flows responsible for the
accumulation of larvae in hydrographic features, if any, and how the mechanism
responds to large-scale disturbances and the mechanism's alongshore variability.
There is evidence that internal tidal bores can transport larvae (Pineda,
1991; Pineda, 1995; Leichter, et al., 1996; Leichter, et al., 1998).
Internal tidal bores in Southern California
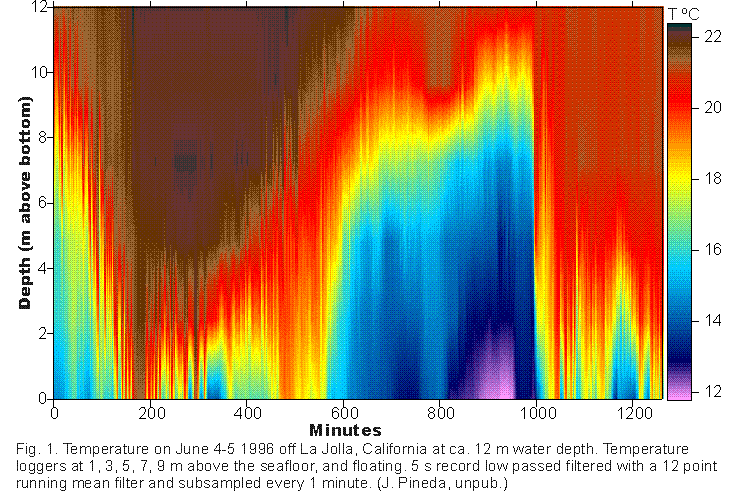
Sharp surges of cold water (Winant, 1974) that often surface the thermocline
at diurnal or semidiurnal periodicities have been identified as internal
tidal bores; (Cairns, 1967; Pineda, 1991; Pineda, 1994) (Fig.1)
Internal tidal bores occur in groups of events lasting 2-10 days in Spring and Summer, when the water column is strongly thermally stratified. Groups of events are unpredictable in the short term. Periodicity in surface water temperature suggests that, on average, internal tidal bores tend to occur around neap tide (Pineda, 1991; Pineda, 1995). During these events, temperature records imply that the nearshore water body is replaced at diurnal or semidiurnal periods for several days. (Figure 2 and 3).
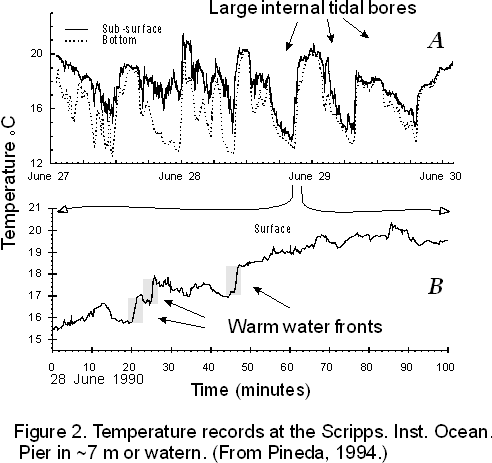
The sequence of events during an internal tidal bore event has been interpreted as follows. At first, a cold water bore is advected shoreward (Cairns, 1967), displacing warm water offshore. This creates an imbalance in hydrostatic pressure between heavier inshore water and lighter offshore water. A front forms between the inshore cold water and the displaced offshore warm water. Then, as cold water recedes offshore because of its density, currents reverse, and a "warm" internal bore returns, pushing the front all the way back to the shore (Figure 3)(Pineda, 1994).
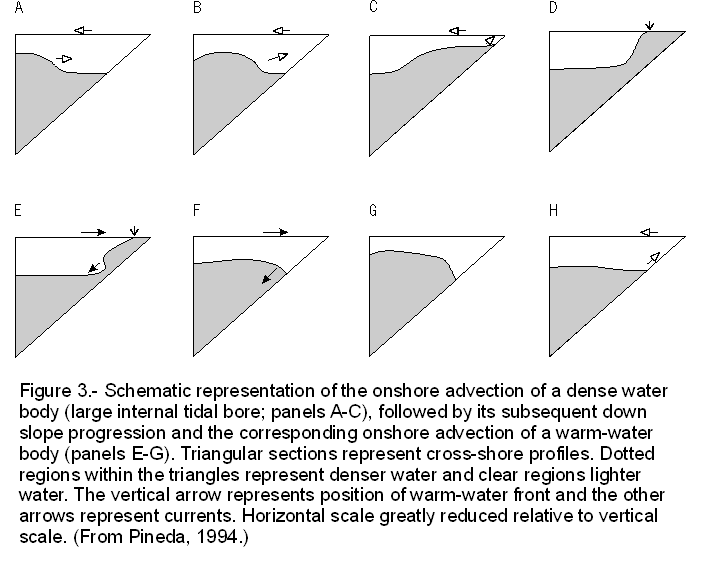
Recent observations on the onshore migration of internal-bore warm-fronts
in shallow water revealed that fronts are followed by large-amplitude high-frequency
internal waves. Warm bore fronts had concentrating currents resulting from
high-frequency internal motions and a larger two-way flow, with surface
onshore currents, and bottom offshore. Onshore currents were faster closer
to the seasurface, while faster offshore currents occurred at a distance
from the bottom. In some cases, averaged onshore currents ![]() >
>![]() , propagation speed.
Faster currents than the rate of advance of the front implies that the
origin of surface material accumulated in the front is behind the fronts,
not
in front of the front. Three nearshore invertebrate larval taxa showed
contrasting horizontal and vertical distributional patterns. All were found
in all sites across fronts, but only larvae of two shore barnacles,
Chthamalus
spp. and Pollicipes polymerus, were concentrated at the front's
surface, while the subtidal bryozoan Membranipora spp.
was not.
Pollicipes
was
more abundant in the fronts' surface, and appeared to be more concentrated
than Chthamalus. Observed frontal downwelling currents suggest that
larvae would have to swim upward to withstand currents and accumulate.
Larval distribution around the front suggests that in some cases frontal
Pollicipes
originates offshore if the front, while
Chthamalus
may originate
at either side of the front.
, propagation speed.
Faster currents than the rate of advance of the front implies that the
origin of surface material accumulated in the front is behind the fronts,
not
in front of the front. Three nearshore invertebrate larval taxa showed
contrasting horizontal and vertical distributional patterns. All were found
in all sites across fronts, but only larvae of two shore barnacles,
Chthamalus
spp. and Pollicipes polymerus, were concentrated at the front's
surface, while the subtidal bryozoan Membranipora spp.
was not.
Pollicipes
was
more abundant in the fronts' surface, and appeared to be more concentrated
than Chthamalus. Observed frontal downwelling currents suggest that
larvae would have to swim upward to withstand currents and accumulate.
Larval distribution around the front suggests that in some cases frontal
Pollicipes
originates offshore if the front, while
Chthamalus
may originate
at either side of the front.
Literature cited
Cairns, J.L. 1967. Asymmetry of internal tidal waves in shallow coastal waters. J. Geophys. Res. 72, 3563-3565.
Leichter, J.J., G. Shellenbarger, S.J. Genoveseet al. 1998. Breaking internal waves on a Florida coral reef: a plankton pump at work? Mar. Ecol. Prog. Ser. 166, 83-97.
Leichter, J.J., S.L. Wing, S.L. Milleret al. 1996. Pulsed delivery of sub-thermocline water to Conch Reef, Florida keys by internal tidal bores. Limnol. Oceanogr. 41, 1490-1501.
Pineda, J. 1991. Predictable upwelling and the shoreward transport of planktonic larvae by internal tidal bores. Science (Washington D.C.). 253, 548-551.
Pineda, J. 1994. Internal tidal bores in the nearshore: warm-water fronts, seaward gravity currents and the onshore transport of neustonic larvae. J. Mar. Res. 52, 427-458.
Pineda, J. 1995. An internal tidal bore regime at nearshore stations along western USA: predictable upwelling within the lunar cycle. Continental Shelf Research. 15, 1023-1041.
Winant, C.D. 1974. Internal surges in coastal waters. Journal of Geophysical Research. 79, 4523-4526.